本稿は水波誠『昆虫──驚異の微小脳』のノートである.
(水波誠,2009,昆虫──驚異の微小脳,中央公論新社,東京.)
第1章 昆虫の繁栄を支える小さな脳
ハエの複雑な体
ハエには鬱陶しいだけの愚かな生き物というイメージがある.
しかしハエの体は複雑な機能と構造を持つ.
- 開放血管系
-
- 昆虫には血管系がなくすべての組織や器官を直接血液に浸されている.
- 気管系
-
- 昆虫の血液には酸素を運ぶヘモグロビンはなく,酸素は気門から気管系によってじかに体の組織に運ばれる.
これは空冷式エンジンに似ている.
小型のエンジンは空冷式の方が水冷式によりも効率が良いのと同様,ハエのように小さな動物では空気を直接組織に運ぶ方が効率が良い.
いらいらさせられる原因
複眼の時間分解能は人の数倍も高く,ハエには蛍光灯が点滅して見える.
またハエ目の昆虫は後ろの2枚の翅が退化して,平均棍と呼ばれる棒状になっており,飛行を安定させるジャイロスコープの役割を果たす.
*
私たちがハエにイライラさせられるのは,ハエが愚かな動物だからではなく,手強いライバルであることを本能的に察知しているからではないかと考えたくなる.
昆虫──陸の王者
実際昆虫の地球上の全種類は全ての動物の2/3を占めており,昆虫は陸の王者と言える.
- 節足動物門→昆虫綱→コウチュウ目,チョウ目,ハチ目 (この3つの目で動物種の半分を占める)
- 古生代シルル紀デボン紀にかけて浅い,海を生活の場としていた甲殻類の一部が陸上に進出し,昆虫の祖先となった.
- 最初に出現した昆虫は翅を持っていなかった.
昆虫はなぜ繁栄したか
昆虫はキチン質の外骨格を持ち,陸上で水分蒸発から身を守れるようになった.
また体が小さいため,食べ物が少なくて済み,住み分けも容易である.
以上は昆虫だけでなく他の節足動物 (ムカデやクモなど) にも当てはまる.
*
昆虫に特有の事情としては次の3つが挙げられる.
- 翅を獲得し,高い移動能力能力を実現したこと.
- 変態によって成長と繁殖の完全分離を実現し,効率的な資源利用を可能にしたこと.
- 幼虫期は食事に専念し,成体になってからは移動と繁殖に専念できる.
- 花をつける植物 (被子植物) と共生関係を結んだこと.
- 昆虫と被子植物は互いに依存しながら爆発的な種分化を起こし,陸上生態系の骨格を作ってきた.
しかしながら以上の飛翔能力,変態の制御,被子植物の認識と記憶において,神経系の働きが重要な役割を担っていることは見過ごされてきた.
微小脳の驚異
昆虫の頭部にある食道上神経節は感覚情報の統合・記憶・運動の制御などを司る (図1参照).
これを微小脳と呼ぶことにする.
人の脳をスーパーコンピューターに例えると,昆虫の脳はせいぜいノートパソコン程度であるが,市場という生存競争の場ではノートパソコンが勝利を収めているのと同様,生態系という生存競争の場では,小型・軽量・低コストの昆虫の微小脳の方が圧倒的に成功していると考えられる.
そこで以下では昆虫の微小脳の「賢さ」について見ていく.
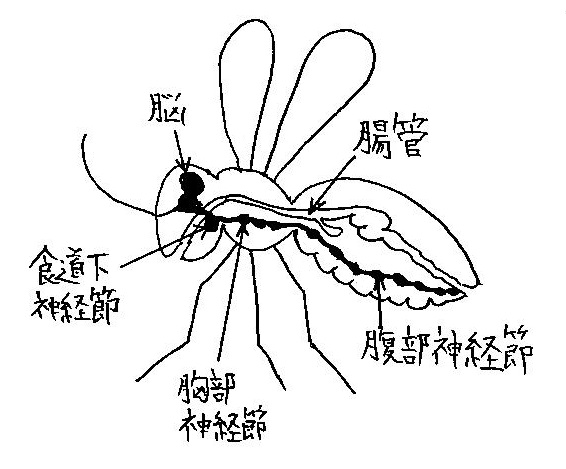
図1 昆虫の神経系
第2章 ファーブルから現代まで
科学的な行動研究の始まり
『ファーブル昆虫記』には昆虫の本能行動が数多く記されている.
例えばジガバチは昨日掘った巣穴の場所を記憶ていて,翌日も同じ場所に戻って来られる.
ファーブルは本能行動についての観察試料を残し,本能研究の先駆者となったが,それらを学問的に体系づけるには至らなかった.
科学的な行動研究はフリッシュ,ローレンツ,ティンバーゲンの研究に始まる.
- カール・フォン・フリッシュ
- 観察と実験により,ミツバチが餌場の場所をダンスにより仲間に伝えていることを確かめた.
- コンラート・ローレンツ
- 動物の行動をありのまま観察することに努め,ハイイロガンは生後一定期間内に見た身の回りの動くものを親とみなして,後追い行動をするようになること[刷り込み]や,動物の本能的な行動を引き起こす鍵刺激の存在を発見した.
- ニコラス・ティンバーゲン
- 実験と観察の両方に優れており,ハチは巣穴近くの景色を頼りに起巣をすることなどを発見した.
現在の神経行動学は,動物が自然界で示す行動の神経機構を明らかにすることを目指している.
行動の仕組みの研究
バッタのジャンプに直接関係するニューロンが同定され,ジャンプの仕組みが比較的少数のニューロンからなる神経回路の働きにより説明できることが明らかになった
ここから「昆虫の行動の仕組みは,単一のニューロンの活動に関連して説明できる」という確信が深まっていった.
しかし昆虫の脳が100万個のニューロンからなることから想像されるように,行動に間接的に関与するニューロンまで含めると,ニューロンの数は多数にのぼり,昆虫の行動はそれほど単純には説明できないことが後になって分かった.
現在の研究の最前線
昆虫の行動研究の基本テーマは,「昆虫の微小脳の仕組みが哺乳類の巨大脳の仕組みとどこまでが共通でありどこからが異なるのか」であると著者は考えている.
- 第3章 複眼
- 第4章 単眼
- 第5章 飛翔
- 第6章 嗅覚
- 第7章,第8章 学習・記憶
- 第7章,第9章,第10章 起巣・コミュニケーション
- 第11章 まとめ
十九世紀のアカデミー論争
神経系は節足動物では腹側に走っているに対し (図1参照),脊椎動物では背側に走っている.
キュヴィエはこのことを根拠に節足動物と脊椎動物を区別した.
これに対しジョフロウは,節足動物と脊椎動物は上下がひっくり返ったにすぎず,本質的には同じボディー・プランであると考えた.
ボディー・プランを支配する遺伝子
中枢神経系の形成に関わる,昆虫と脊椎動物に共通の遺伝子群があり,そのような脊椎動物の遺伝子を昆虫に入るとそこに神経系が形成されることから,現在ではキュヴィエとジョフロウの論争は,ジョフロウが正しかったと見られている.
[これは脊椎動物に関して言えば,背側に神経系ができるというよりもむしろ神経系の形成される方が背側になるのであり,「腹側」「背側」という概念は体内の組織との位置関係と無関係にあるものではないと理解すれば良いだろうか.]
またこのことは,昆虫と脊椎動物の脳に共通する機能が必ずしも独立に進化してきたものではなく,その基本的な部分は共通の祖先から引き継いだものであることを示唆している.
神経系の初期発生
動物は進化の過程で,昆虫に繋がる旧口動物と,脊椎動物に繋がる新口動物に分かれた.
旧口動物では口から肛門に向かって消化管が形成されるに対し,新口動物では肛門から口に向かって消化管が形成される.
*
昆虫については体節と呼ばれる繰り返し構造が生じ,体節ごとに1対の足と1つの神経節が形成される (図1参照).
体節ごとの機能の特殊化
成長が進むと前の6つの体節は頭部になり,いくつかの脚は触覚や口器になる.
前方の3つの体節の神経節が融合して脳が作られる (図2参照).
- 前大脳 (連合中枢) + 視葉 (視覚情報の処理)
- 中大脳 (嗅覚・接触感覚・風感覚の処理)
- 後大脳 (上あごの接触感覚・頭部の感覚毛からの風感覚情報の処理)
前大脳 (特にキノコ体) は並列的な,場合によっては異なる行動を要求するような感覚情報を統合して,1つの行動を選択するような連合中枢であり,これが発達したことで,個体としてのまとまった行動が可能となったと考えられる.
- 食道下神経節 (味覚や接触運動の制御) ↔ 脊椎動物の延髄
- 胸部神経節 (歩行や飛翔の制御),腹部神経節 (呼吸,消化,排泄,交尾,産卵等の制御) ↔ 脊椎動物の脊髄

図2 前大脳・中大脳・後大脳
第3章 複眼は昆虫の何をものがたるか
二つのタイプの眼
ヒトの眼はレンズ眼であるのに対し,昆虫は六角形の個眼が敷き詰められた複眼を持つ.
- トンボの複眼の背側 (上方) には茶色の斜光色素があり,サングラスの役割をする.
- ハナアブの雄は左右の複眼が正面で接しているのに対し,雌の左右の複眼は離れている.これは交尾の際,雄が雌を正面に捉えて追尾するという非対称性を反映している.
眼の進化
脊椎動物のレンズ眼はオウムガイなどが持つピンホール眼に1枚のレンズが付け加わる形で進化したと考えられる.
一方,節足動物の複眼は,ゴカイなどの環形動物が持つ色素細胞を備えた管の各々にレンズが付け加わる形で進化したと考えられる (図3参照).
*
光受容細胞の光受容部位は,旧口動物では微絨毛であるのに対し,新口動物では繊毛が変形したものである.
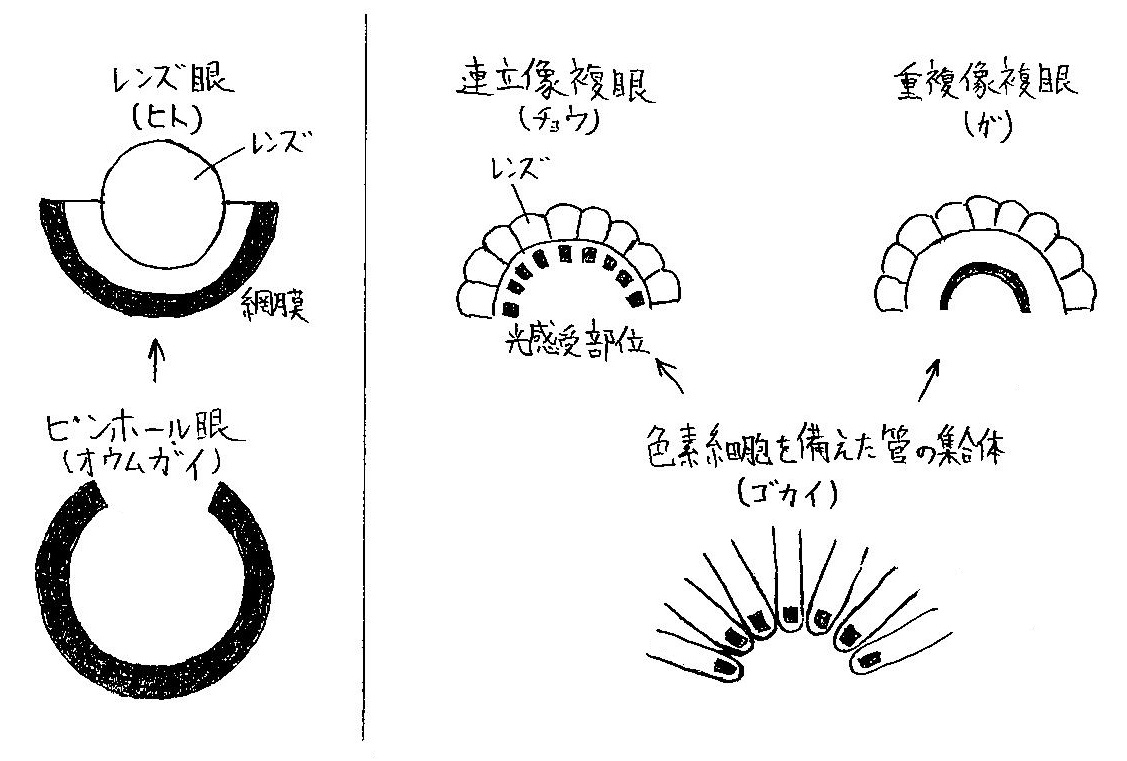
図3 さまざまなタイプの眼
画素数一万二〇〇〇の視覚世界
例えばイエバエの複眼は左右合わせて1万2千個の個眼を持つ.
これをデジカメの画素数 (例えば500万) と比べると,昆虫には粗い像しか見えていないことが分かる.
ヒトと比べても,昆虫の視力は2桁も低い.
*
複眼の解像度を上げるには,レンズを小さくして個眼を増やせば良い.
ところがレンズを小さくすると光の回折が著しくなり,鮮明な像が得られなくなるというジレンマがある.
このような洞察に基づきキルシュフェルトは,ヒトと同じ解像度を持つ複眼を得るには,直径1mの巨大な複眼を要すると結論した.
【参考】
Feynmanはミツバチの個眼が実際にほぼ最適なサイズとなっていることを示している.
複眼の半径を  ,個眼のレンズの直径を
,個眼のレンズの直径を  とすると,レンズを小さくするにつれて隣り合う個眼の間の角度は
とすると,レンズを小さくするにつれて隣り合う個眼の間の角度は  に従って減少する (すなわち解像度が増す) のに対し,レンズに入る光 (波長
に従って減少する (すなわち解像度が増す) のに対し,レンズに入る光 (波長  ) の回折による開きの角度は
) の回折による開きの角度は  に従って増大する.
に従って増大する.
そこでレンズの最適なサイズ を,和  が最小になるような の値
が最小になるような の値  と考えると,これは
と考えると,これは  に対して
に対して  を与え,文献値 30μm とよく一致する.
を与え,文献値 30μm とよく一致する.
(ファインマンほか,2012,ファインマン物理学 II(富山小太郎訳),株式会社岩波書店,東京,pp.141–142.)
*
しかし昆虫のように小さな動物はそれだけ近くのものだけを見分けられれば十分であり,このことを反映して,動物の視力は大まかには,体高が1/10に減少すると1/10だけ減少する.
またハエなどの昆虫は複眼の空間的解像度の低さを,時間分解能の高さで補っている.
二つのタイプの複眼
連立像眼は感桿 (光受容部位) がレンズの直下にあり,その個眼からの光しか受容しないためシャープな像が得られ,昼行性の昆虫に多く見られるのに対し,重複像眼は感桿 (光受容部位) がレンズから離れて位置し,真上のレンズだけでなく周囲の個眼のレンズからの光も受容するため光を効率良く集められ,夜行性の昆虫に多く見られる (図3参照).
- 視野の広さ:複眼では個眼を並べれば視野を広げられる.イトトンボの視野は全立体角をカバーしている.
- 時間分解能:ハエの複眼は時間分解能が高く,映画のコマや蛍光灯のちらつきを識別できる.
- これはハエの時間感覚がヒトとは異なり,人から見れば短命なハエも主観的には長生きしていることを意味しているのかもしれない.[ハエに主観があるならば.動物が経験する,動物に固有の主観的な世界は環世界と呼ばれる.]
- ハエの複眼の時間分解能の高さは,複眼という眼の構造によるものではなく,光受容の化学反応の速さに由来する.
- 明暗変化の検知:視覚系のニューロンは一定の光を受け続けると順応するため,物が見えなくなる.
- ヒトは眼球を常に動かして順応を防いでいる.[サッカード]
- 昆虫は止まっているときには,ゆっくり忍び寄る敵が見えず,一方移動中は景色は相対的に動いているため見えている.
色を見る
- フリッシュはミツバチが色を見分けていることを実験的に確かめた.
- 多くの昆虫は紫外,青,緑に反応する視細胞を持ち,これらが昆虫の3原色を成す.これはヒトの場合の3原色 (青,緑,赤) に比べて短波長側にずれている.
- ミツバチでは例えば青緑と紫外線を混ぜると,白の感覚を与えることが行動実験から分かっており,青緑と紫外線は補色となっていると考えられる.ミツバチには紫外線と緑の光に興奮性を示し,青の光に抑制性を示し,それらを混ぜた「白」の光には反応しない色対比ニューロンがあり,これが反対色への反応に関わっていると考えられる.
- 昆虫は花の中央の紫外線を吸収する領域に引きつけられる.(花びらの向きを逆にすると,昆虫は花びらの外側に向かう.)
- ヒトには,青色の紙はどのような波長の光で照らしても青色に見える.これを色の恒常性と呼ぶ.色の恒常性はアゲハチョウにもあることが実験的に分かった.
【参考】
人間の網膜には赤・緑・青に対応する色細胞があることは確かに,人間が見る世界にとっての三原色が赤・緑・青であることの説明にはなっているかもしれない.
しかし任意の色 X が X=aA+bB+cC と表されるような三原色 A,B,C は,係数 a,b,c が負の値をとることも認めれば,赤・緑・青に限らず任意に選ぶことができる.
ただし赤・緑・青の組合せは,負の係数を用いずに多くの色を実現できるような組合せではある.
(ファインマンほか,2012,ファインマン物理学 II(富山小太郎訳),株式会社岩波書店,東京,pp.123–124.)
ラモニ・カハールのためいき
カハールはゴルジ染色法を用いて,神経がニューロンから成ることを見出した.
カハールは昆虫の視葉を観察し,その高等ほ乳類の網膜にも劣らない精巧な構造に驚いている.
(視葉に限らず,「昆虫の微小脳は小さなスペースに多くの機能を整然と詰め込んだ『集積回路』」と言える.)
視覚中枢の構造と働き
視覚情報は視葉 (視覚中枢) を構成するラミナ,メダラ,ロビュラ複合体に順次送られる (図4参照).
その際,網膜での視覚情報の配列を1対1に写し取るように情報が伝えられる (視野再現構造).
同様の構造は哺乳類の大脳視覚野でも見られる.
- ラミナ:明暗のコントラストを強調
- 光が当たる領域の周囲ではニューロンの応答が抑制されること (側抑制) による.これは昆虫に限らず,動物の眼の基本原理.
- メダラ:物体の局所的な運動を検出
- 各個眼から隣接する個眼に向かう6つの方向について,順方向に物体が動くと興奮し,逆方向に物体が動くと抑制性を示す,局所運動検知ニューロンがある.
- ロビュラ複合体 (ロビュラ+ロビュラプレート):空間の大域的な情報を抽出

図4 視葉 (視覚中枢) の構造
動きの方向の神経計算
ハッセンシュタインとライヒャルトは,メダラおよびロビュラプレートでの運動検知を次のように定式化した.
すなわち隣接する空間領域A,Bからの視覚情報  は,時間
は,時間  だけ前の信号
だけ前の信号  と掛け合わされて出力
と掛け合わされて出力

となる.
このとき簡単のために図5のような入力を考えると,出力は
- 物体がB→A方向 (順方向) に横切ったとき,正の応答 (+1)
- 物体がA→B方向 (順方向) に横切ったとき,負の応答 (-1)
- A,Bに同時に物体の像が現れたとき,応答なし (0)
となっている.

図5 運動検知の神経計算モデル
実験的にもハエに対して,隣接する空間領域に向けられた2つの視細胞を刺激し,その信号を受け取るHIニューロンと呼ばれるロビュラプレートの大型のニューロンの応答を調べたところ,モデルと同様の振舞いが見られた.
【コメント】
このモデルは神経機構に関する生理学的な知見からボトムアップ的に導かれるものではなく,むしろ生理学的な知見を踏まえつつも,神経系の働きをトップダウン的に解釈するものであると言えるだろう.
オプティカルフロー (画像の流れ) を見る
運動すると周りの景色が相対的に動く.
この景色の流れをオプティカルフローと呼ぶ.
ハエのロビュラプレートには,小さい物体の動きにはあまり反応せず,視野全体にわたるオプティカルフローに反応するニューロンがあることが知られている.
形の識別
Y字型の分かれ道の一方に餌を置き,2本の道それぞれのの奥に異なる図形を提示して,ミツバチに正しい図形 (餌のある方の図形) を学習させた.
傾きの異なる線分を2つの図形に用いたところ,ミツバチは正しい傾きの線分を選ぶことができた.
ミツバチの錯視
それだけでなく,図6のような「カニッツァの図形」を用いた場合にも,ミツバチは実在しない長方形を識別して,その傾きの正しい方を選択した.
ミツバチがヒトと同じ錯視を示すことは,視覚系の情報処理の類似性を示唆する.
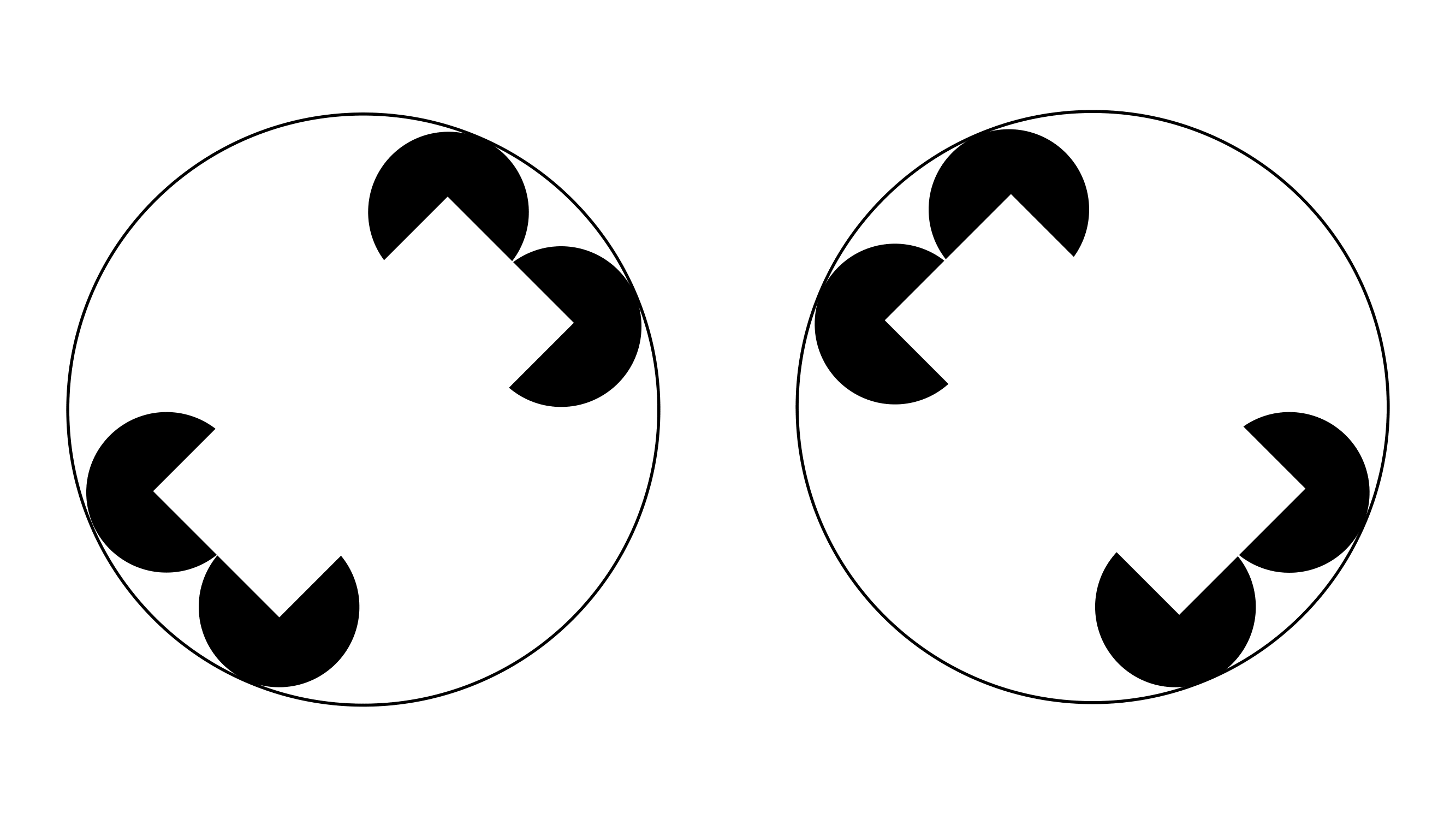
図6 カニッツァの図形
高度な視覚認知
また対称な (または非対称な) 図形を選ぶようにミツバチを訓練すると,テストの際に訓練で用いたのと異なる図形を提示しても,ミツバチは対称な (または非対称な) 方の図形を選ぶことができた.
さらに分岐の入り口で見本となる色を見せ,Y字通路の奥の見本と同じ (または異なる) 色を選ぶように訓練したミツバチは,入り口で色だけでなく図形や匂いを提示した場合にも,見本と同じ (または異なる) 図形や匂いを選ぶことができた.
このことからミツバチには,対称性と非対称性を区別して一般化する能力や,ものの(非)同一性という抽象的な概念を抽出し,他の感覚に転移する能力があることが分かる.
第4章 単眼はどんな働きをしているか
第三の眼
多くの昆虫は3個の単眼を持つ.
脊椎動物の祖先にも,額に額眼 (中央眼) と呼ばれる第3の眼を持つものが多くいた.
単眼の構造と機能的な特徴
単眼は明暗の検知しかできない単純な光受容器である.
- 単眼のレンズは光受容細胞層のはるか後方に焦点を結ぶ.[ピントが合っていない.]
- 光受容細胞の情報を受け取る2次ニューロンは数十個程度しかない.
- 単眼の情報は少数のシナプスを介して胸部神経節の運動中枢に送られ,複眼の情報よりも素早く伝達される.
→ 単眼は像の知覚には向いていないが,空間解像度を犠牲にして明暗変化を感度良く検知することに特化しており,粗い情報を基にした素早い反応に関係している.
単眼の行動上の役割
単眼で受容した明るさの情報は,明け方の行動の開始や夕暮れの行動の終了を決定するのに用いられる.
*
単眼は空と地面の明るさのコントラストを検知し,飛翔姿勢を制御するのに役立てられているとウィルソンは考えた.
すなわち昆虫の飛翔中に体が傾くと,3つの単眼の明暗は体の傾き方に応じた変化をするため,姿勢の変化を検知できる.
この仮説はバッタやトンボの行動実験によって検証された.
(なお昆虫の単眼による飛翔制御は,単眼を模した3個の光センサーを搭載したラジコン飛行機などに応用されている.)
単眼は単純であるがゆえに素早く感度良く明暗を検知できる「simple is best」なデザインとなっており,「simple is best」は単眼だけでなく昆虫の微小脳の本質でもあるかもしれない.
単眼系の神経回路の多様性
「大学では,現代生物学の力点が,眼の前にいる個別の生き物を理解することではなく,むしろ多様な生き物に共通する一般原理を探ることにあることを学び,一種のカルチャーショックを受けた」(p.82) とあるのが面白い.
*
ワモンゴキブリは繁殖力が高く,飼育が容易であり,微小電極法を用いて脳のニューロンの活動を記録できるため,神経系の研究に適している.
*
- 3シナプス回路 (例:ワモンゴキブリ)
- 単眼の光受容細胞 → 2次ニューロン (少数) → 3次ニューロン → 脳内の諸中枢
- 視覚情報の,少数の2次ニューロンへの収斂 → 明暗検知の感度が良くなる (感度重視型)
- 2シナプス回路 (例:ミツバチ)
- 単眼の光受容細胞 → 2次ニューロン → 脳内の諸中枢
- 少数のニューロンへの情報の収斂を省略し,シナプス数を減らす → 信号伝達が速くなる (速度重視型)
- 複合型 (例:トノサマバッタ)
- 2シナプス回路と3シナプス回路の両方を持つ.
単眼系の神経回路の進化
進化の過程で生活様式の変化が起こっても,このような神経回路の配線タイプの違い (感度/速度重視型) を決める,神経発生プログラムの変化は起こりにくかったと考えられる.
例えばヤガは夜行性であるため,感度重視型の配線の方が有利であるにも関わらず,実際には速度重視型の神経回路を持っているのはこのためであると考えられる.
単眼系を用いた明暗視の神経機構の研究
昆虫に限らず多くの視覚系では,光受容細胞から2次ニューロンへの情報伝達の際,明暗コントラストの情報が抽出される.
物体の反射率は決まっているため,照明光の強度が変化しても物体と背景のコントラストは一定であり,動物の視覚はこの明暗コントラストを用いることで物体の特徴を捉えることを可能にしていると考えられる.
そして実験的に調べやすい昆虫の単眼系の研究を通して,明暗コントラストの抽出に関わる神経機構の一般原理を明らかにできるかもしれない.
単眼の光受容細胞での明暗情報の符号化
入力光 (平均強度  とその周りの変化 (変調)
とその周りの変化 (変調)  の和)
の和)
↓
光受容細胞の応答 (膜電位の変化, に対応する直流成分  と に対応する変調成分
と に対応する変調成分  )
)
↓
2次ニューロンの応答 
↓
3次ニューロンの応答 
光受容細胞の応答  は光強度
は光強度  の対数に比例することが分かった.
の対数に比例することが分かった.
このとき応答の変調成分 は近似的に明暗コントラスト  に比例することになり,コントラストを符号化していると言える.
に比例することになり,コントラストを符号化していると言える.
【考察】
光強度と光受容細胞の応答の間の対数的な関係

(定数  は真数を無次元化するのに必要とされる) を仮定し,2式を辺々引くと
は真数を無次元化するのに必要とされる) を仮定し,2式を辺々引くと

となる.
よって  を仮定しその1次までとると,確かに変調成分 と明暗コントラスト の比例関係
を仮定しその1次までとると,確かに変調成分 と明暗コントラスト の比例関係

が得られる.
【コメント】
光受容細胞の応答が明暗コントラストを符号化していることは,応答の光強度の対数に比例していることから説明されるが,応答の光強度に関する対数的な振舞いを神経機構に還元して説明するには至っていない.
単眼の光受容細胞と二次ニューロンの間のシナプスでの信号処理
2次ニューロンの応答 では,光受容細胞の応答の直流成分 が圧縮され[捨てられ],明暗コントラストを符号化した が“取り出される”.
ただし光受容細胞と2次ニューロンをつなぐシナプスの性質上, と の符号は逆転する.
単眼二次ニューロンの応答特性
[以上により2次ニューロンの応答 は明暗コントラスト を検知したものとなっているはずである.実際]
2次ニューロンの応答 は明暗コントラスト に比例することが,実験により直接的に確かめられる.
なお強度  の刺激が突然
の刺激が突然  だけ変化したときに,心理的に感じられる強度変化がコントラスト
だけ変化したときに,心理的に感じられる強度変化がコントラスト  に比例することはウェーバーの法則と呼ばれ,このことは2次ニューロンの応答がウェーバーの法則に従うことを意味している.
に比例することはウェーバーの法則と呼ばれ,このことは2次ニューロンの応答がウェーバーの法則に従うことを意味している.
単眼二次ニューロンと三次ニューロンの間のシナプスでの信号処理
3次ニューロンは,明るさの平均レベルからの減少に対する2次ニューロンの正の極性の応答を増幅し,明るさの増加に対する2次ニューロンの負の極性の応答をほとんどゼロにするような,「半波整流器」の役割をする.
明るさの増大よりも減少に大きく反応することは,陰影を検知し,飛翔中に地面に突っ込むのを防ぐことや,外敵に気付く意味があると考えられる.
*
以上を改めてまとめると,単眼系の情報処理は次のようになる.
光入力
↓ 光受容細胞 (対数変換器)
光強度の対数の応答
↓ 2次ニューロンに繋がるシナプス (直流応答の除去)
コントラストに比例する応答
↓ 3次ニューロンに繋がるシナプス (整流器)
減光,陰影への応答
明暗検知回路から運動検知回路への進化
ある種の昆虫の3次ニューロンには,前節で紹介された「暗」の情報を選択的に伝えるオフチャネルだけでなく,「明」の情報を選択的に伝えるオンチャネルや,明るさの増大と減少の両方に正の極性の応答を示す「全波整流器」のオン-オフチャネルを併せ持つ明暗検知系がある.
「全波整流器」は「明」「暗」それぞれに対する「半波整流器」を用いて再現・説明できる.
そして隣接する明暗検知系の間に時間遅れを伴う減算・加算を仮定すると (シナプス整流モデル),運動検知のモデルであるハッセンシュタインとライヒャルトのモデル (第3章) の応答を忠実に再現できる.
これは昆虫に限らず一般に,動物の神経回路は明暗検知のような単純な視覚機能の神経回路が改変されて,運動検知能力を獲得した可能性を示唆している.
第5章 空を飛ぶしくみ
空への進出
昆虫の飛翔の力学や制御の仕組みにはまだ分かっていないことが多い.
昆虫の飛翔の制御系の特徴は「素早さ」であると言える.
昆虫の飛翔のしくみ
本章では昆虫の飛翔について,
- 推進力を安定的に得る仕組み
- 外乱に抗して飛翔姿勢を保つ仕組み
に関係する反射的な制御に焦点を当てる.
反射的な制御は“随意的な”仕組みに制御されており,このような運動制御系の階層構造は哺乳類にも共通している.
このような類似構造はそれぞれ独立に進化したと考えられる (進化的収斂).
バッタの飛翔解析
バッタの飛翔に関する研究から,
- 羽ばたきは周期的であり (1秒に約20回),位相は後翅の方が前翅よりも約7ミリ秒先行すること
- 翅は上下だけでなく前後にも動き,羽ばたきの1周期中に翅の仰角は時々刻々と変化していること
が分かった.
飛翔系の構築
バッタの飛翔について,翅が打ち上げられると打ち下ろし筋が発火し,翅が打ち下ろされると打ち上げ筋が発火する.
発火のタイミングは前翅より後翅の方がわずかに速く,これが羽ばたきの位相差を生み出している.
*
飛翔の基本となるのは,胸部神経節の運動ニューロン・介在ニューロンと脳である.
*
打ち上げ筋・打ち下ろし筋の各々は,わずか2,3個の運動ニューロンの2,3発の活動電位によって収縮する.
運動ニューロンは周期的に発火する.
飛翔系における中枢パターン発生器の発見
バッタの飛翔における周期的な運動パターンを説明する最初の仮説は,感覚フィードバックに基づくものであった.
これは翅が打ち上げられると,その感覚のフィードバックが翅の打ち下ろしを引き起こし,その感覚のフィードバックが翅の打ち上げを引き起こすというような連鎖反応を想定するものである.
ところが感覚フィードバックを断ち切っても,バッタは通常の飛翔パターンを示したため,このような周期的な運動パターンは中枢神経系の振動的な回路 (中枢パターン発生器) によって形成されているものと考えられる.
中枢パターン発生器のニューロン構築
周期的な活動を示すあるニューロンがパターン発生に関与しているのか,それともパターン発生回路から入力を受け取っているだけなのかを判定するのに,「リセット実験」が有用である.
これは注目しているニューロンに強制的に活動電位を発生させ,中枢パターン発生器のリズムがリセットされるか否かを調べる手法である.
このような実験により,バッタの飛翔パターンの発生に関わる個々のニューロンが同定された.
さらにそれらの間のシナプス接続も調べ上げられた.
*
IN301とIN501と呼ばれる2つの介在ニューロンが相互に接続した回路が,飛翔パターンの生成にとって重要であることが実験的に分かっている (INは介在ニューロンを表す).
IN301は打ち上げ期に興奮性入力を受けて,活動電位を発生させる.
IN501は遅れて打ち下ろし期にこの活動電位を受け取り,抑制性シナプス電位を発生させる.
これは直ちにIN301に伝えられ,IN301の活動が抑制される.
するとIN501の発火が停止し,IN301の抑制が解除され,次の発火が可能となる.
これがリズム形成機構の一部を成している.
自己受容器の役割
周期的な運動パターンは感覚フィードバックではなく中枢パターン発生器によると述べたが,感覚フィードバックが羽ばたきに全く無関係というわけではない.
バッタの翅の1枚をある範囲の振動数で強制的に羽ばたかせたところ,その感覚フィードバックにより他の3枚の翅も同調して羽ばたいた.
飛翔姿勢の制御
単眼,複眼,風受容感覚毛の情報は
- 遅く持続的に活動する経路
- 素早く一過的に動作する経路
の2つの感覚運動経路により,飛翔姿勢の修正に用いられる.
以下では後者を取り上げる.
偏差検知ニューロン
単眼,複眼,風受容器からの入力を統合し,ある特定の方向の姿勢の乱れについての情報を選択的に抽出するような,「偏差検知ニューロン」が見つかった.
例えば右側の単眼からの入力を受け,右側の神経索を下降する同側偏差検知ニューロンは,ロール軸 (図7参照) 周りの回転運動が自然な水平の飛翔姿勢から時計回りにずれている場合にのみ反応する.
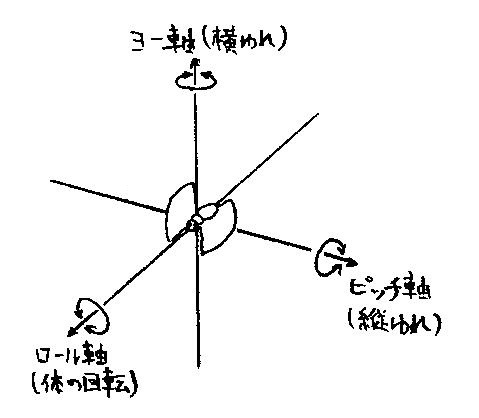
図7 ロール軸,ピッチ軸,ヨー軸
【参考】
ロール軸,ピッチ軸,ヨー軸はいずれも物体 (今の場合,昆虫) に固定した座標軸である.
その空間固定系に対する向きを指定するには3つのパラメータが必要であり,そのようなパラメーターにEuler角を用いることができる.
しかしながらこれは第1の回転 (回転角  ) と第3の回転 (回転角
) と第3の回転 (回転角  ) をいずれも物体固定系の
) をいずれも物体固定系の  軸の周りに行うものであるため,第2の回転で 軸の方向がほとんど変化しないときには と を区別するのが困難となり,実用に向かないという欠点がある.
軸の周りに行うものであるため,第2の回転で 軸の方向がほとんど変化しないときには と を区別するのが困難となり,実用に向かないという欠点がある.
そこで飛行物体を扱う応用分野では,3回の回転を全て物体固定系の異なる軸の周りに行う規約が用いられる.
はじめに機体が水平面内にある場合を考え,機体の軸を  軸にとり,水平面内で 軸に垂直な方向 (翼・翅の方向) を
軸にとり,水平面内で 軸に垂直な方向 (翼・翅の方向) を  軸にとり,鉛直上向きに 軸をとって,
軸にとり,鉛直上向きに 軸をとって, 軸を物体に固定する.
軸を物体に固定する.
まず 軸の周りに角度 だけ回転し,次いで 軸の周りに角度  だけ回転し,最後に 軸の周りに角度 だけ回転する.
だけ回転し,最後に 軸の周りに角度 だけ回転する.
このとき は偏揺れ (yaw) の角, は縦揺れ (pitch) の角, は傾斜あるいはロール (roll) の角を表すことになり,Tait-Bryan角と呼ばれる.
(H.Goldstein他,2008,古典力学(上)(矢野忠ほか訳),吉岡書店,京都,pp.203–204,9.449.)
飛翔制御回路における偏差信号の処理
偏差検知ニューロンからの偏差信号は胸部介在ニューロンに伝達される.
胸部介在ニューロンは中枢パターン発生器からも周期的なシナプス入力を受けており,偏差検知ニューロンの入力がそのタイミングに一致したときだけ飛翔介在ニューロンに信号を送る.
これによって羽ばたきの偏差を適切なタイミングで補正することができる.
*
ニューロン数の少ない昆虫であっても,行動の仕組みを個々のニューロンに還元して説明することは容易ではないが,それは微小脳の理解には不可欠であり,昆虫の飛翔の研究ではそれがある程度成功しているのである.
第6章 匂いを感じるしくみ
嗅覚が果たす役割
- 匂いは食べ物の発見に役立つ
- 植物の香りと昆虫の嗅覚の共進化
- 匂いは異性を引きつける手掛かりとなる (性フェロモン)
- 雄のカイコガは触覚に数分子当たっただけで性フェロモンを検知できる
- 匂いは社会性昆虫の個体間の信号伝達に用いられる
- アリやハチの警戒フェロモン
- 直接触れることで巣仲間を判別する,アリの体表の不揮発性のフェロモン
優れた匂い識別能力
フリッシュはミツバチが複数の匂いの中から特定の匂いを識別・選択できることを実験的に確かめた.
フリッシュの下で学んだ桑原は,ミツバチに匂いと吻伸展反射を関連付けて学習させることに成功し,昆虫でも古典的条件付けが起こることを初めて示した.
ベレシは2種類の匂いを識別させる実験により,ミツバチが無数ともいえる匂いを識別できることを示した.
ワモンゴキブリも各種アルコールとその混合物の匂いを識別することができ,ヒトに匹敵する匂い識別能力を持つ.
ゴキブリやミツバチのような昆虫は,暗闇の中でも触角からの匂いの情報を手掛かりに,花や食べ物の形を識別できると想像されているが,これを証明する実験はまだ行われていない.
嗅細胞,嗅受容体と嗅受容体遺伝子
昆虫の触角には嗅感覚子が分布しており,壁面の小孔から侵入した匂い分子は液体に溶け込み,感覚繊毛の表面の受容体タンパク質と結合する (図8参照).
1個の嗅細胞は1種類の嗅受容体分子を持つため,細胞ごとに特有の匂い応答特性を示す.
これはマウスでも同様であるが,マウスが約1000種類の嗅受容体を持つのに対し,ハエは約60種類の嗅受容体しか持たない.
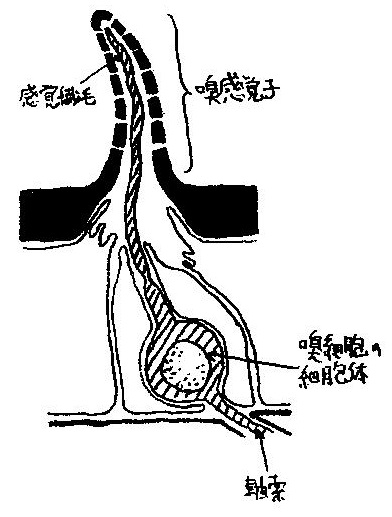
図8 昆虫の嗅感覚子
嗅細胞での匂いの符号化
昆虫でもマウスでも,
- 匂い一般を検知するシステム
- 性フェロモンを検知するシステム
の区別がある.
まずは匂い一般を検知するシステムを見ていく.
*
自然界の匂いは単一の化学物質の匂いではなく,しばしば複数の化学物質の混合物の匂いである.
ゴキブリの嗅細胞の応答を調べると,1個の嗅細胞は1種類の嗅受容体分子しか持たないにも関わらず,複数の匂い分子に反応することが分かった.
また1つの匂い分子は複数の嗅細胞の反応を引き起こすことが分かった.
このことから匂いは多数の嗅細胞の応答パターンによって符号化されていると結論付けられる.
嗅覚中枢での匂いの情報処理
嗅細胞の軸策は触角葉と呼ばれる1次嗅覚中枢に投射して球状の構造 (糸球体) を作り,介在ニューロンとシナプス接続する.
介在ニューロンには,
- 前大脳の側角とキノコ体に投射する投射ニューロン
- 神経突起が触角葉内に限定される局所介在ニューロン
の2種類がある (図9参照).

図9 ワモンゴキブリの中枢嗅覚経路
異なる嗅受容体を持つ嗅細胞の軸策は異なる糸球体に投射するため,触角葉では匂いは,糸球体を単位として,空間的に配列した複数の糸球体の,匂いに応じた応答パターンとして符号化されていると考えられている.
同じことは哺乳類の嗅球内の糸球体にも当てはまり,また哺乳類も昆虫の投射ニューロン・局所介在ニューロンに対応するニューロンを持つ.
このような類似構造はそれぞれ独立に進化したと考えられる (進化的収斂).
*
触角葉の入力ニューロン (嗅細胞) よりも出力ニューロン (投射ニューロン) の方が,狭い範囲の匂いの刺激に応答することが分かった.
これは局所介在ニューロンの側抑制により応答コントラストが強調されるためであると考えられ (視覚系での像の輪郭の強調と同様の仕組み),哺乳類の嗅球にも同様の働きが見られる.
嗅覚中枢での匂いの時間的符号化
投射ニューロンの同期的な発火を阻害すると,ミツバチは構造の似た分子の匂いを識別できなくなることから,匂いの情報は投射ニューロンの同期的な発火という時間的なパターンにも含まれていると考えられる.
*
投射ニューロンから受け取った匂いの情報を前大脳の2次嗅覚中枢がどのように読み出しているのかは,まだよく分かっていない.
性フェロモンの発生源への定位行動
次に性フェロモンの検知の仕組みを見ていく.
匂い物質は匂いの源から遠ざかるにつれて濃度が連続的に低下するような分布にはなっておらず,実は小さい多数の断続的な塊となって浮遊していることが分かっており,このため匂いの源にたどり着くには,濃度の高い方へ進んでいけば良いというわけではない.
*
ガはフェロモンの匂いを検出すると風上に向かって進み,匂いを見失うとターンして匂いを含む風の中に戻る.
これを繰り返すことで,ガは風上に向かってジグザグ定位する.
同様のジグザグ定位は魚や鳥,エビなどの匂い源探索行動にも見られる.
*
カイコガは体が重くて飛ぶことができない.
そこでカイコガの雄に1回だけフェロモン刺激を与えて歩行パターンを調べると,初めは刺激を受けた側に進み (サージ),次いで蛇行 (ジグザグターン) を数秒間行い,ループに移行することが分かった.
よってフェロモン刺激と高頻度で出会う場合には,サージやジグザグターンの段階で刺激に出会ってプログラムがリセットされるため,ほぼ直線的な歩行パターンが実現されるのに対し,フェロモン刺激が低頻度となると,複雑なループ軌道をとることが説明できる.
性フェロモン情報の脳内回路
性フェロモンを受容する嗅細胞は一般の匂いの受容細胞と違って,特定の性フェロモンの匂いにのみ鋭く反応する.
*
性フェロモン受容細胞の軸策は大糸球体に収斂し,大糸球体のニューロンは前大脳の側角とキノコ体に投射する.
ただし性フェロモンの情報を受け取る側角の領域は,一般の匂いの情報を受け取る領域とは異なっている.
側角とキノコ体のニューロンは前大脳の側副葉などに出力する.
匂い源への定位行動に関する運動指令は,主に側副葉から下降性ニューロンによって胸部神経節の運動中枢に伝えられる.
*
カイコガは性フェロモンの刺激を受けるたびに興奮状態と静止状態が入れ替わる下降性ニューロンを持つ.
このような応答はフリップ・フロップ応答と呼ばれる.
このフリップ・フロップ神経活動は首の向きを変えるターンを促すことが分かっており,左右の側副葉間の抑制性の連絡によって生み出されると考えられている.
匂い源を探索するロボット
本物のカイコガの触角をセンサーに用い,得られた電気信号をフリップ・フロップ型のプログラムで処理する,カイコガの匂い源探索の戦略と神経機構を模倣したロボットを作成し,カイコガの匂い源探索行動を再現した研究がある.
湿度や温度を受容する細胞
昆虫は温度や湿度の感覚が発達している.
体の小さい昆虫は体の体積に対して表面積が相対的に大きいので,外界の湿度や温度の影響を受けやすい[体のサイズaに対して(表面積)/(体積)~1/a].
ミツバチは変温動物であるが,働きバチが翅で風を送り込んだり巣に水を持ち運んだりして巣の温度を一定に保つため,コロニー全体では“恒温動物”として振舞う.
*
ワモンゴキブリはキノコ状の幹構造を有する感覚子を持ち,その中には
- 湿受容細胞
- 乾受容細胞
- 冷受容細胞
が入っている.
湿受容細胞と乾受容細胞はそれぞれ相対湿度の上昇と低下に反応する.
(相対湿度とは水蒸気量の飽和水蒸気量に対する割合であり,飽和水蒸気量は温度とともに増加する.)
これらは幹構造の吸湿に伴う体積変化を機械的変位として捉えているという仮説が有力である.
ワモンゴキブリの場合,冷受容細胞は湿度・温度受容感覚子だけでなく,脂肪酸に反応する嗅細胞の感覚子の中にも入っている.
「温」受容細胞はゴキブリの触角では見つかっていない.
*
ヒトは湿度を皮膚の温度や触角などの複合感覚と捉えており,湿度専門の受容器を持つ昆虫にある意味では劣っていると言える.
一方,ヒトの皮膚には温度専門の受容細胞があり,温度を体性感覚として捉えているのに対し,昆虫の触角は空気中の温度勾配を捉えて遠くの温度環境を感知する,言わば「リモートセンシング感覚」である.
触角葉の温度・湿度感覚糸球体
嗅細胞と同様,湿・乾・冷受細胞の軸策末端は,触角葉においてそれぞれ湿糸球体・乾糸球体・冷糸球体を形成することが分かった.
これらが機械感覚の1次中枢である中大脳背側葉と隣接していることは,温度と湿度の感覚細胞が機械受容細胞から進化し,リモートセンシング感覚である嗅覚と同様に触角葉で情報処理を行うようになったことを示唆している.
前大脳の温度・湿度感覚中枢の発見
触角葉の湿,乾,冷糸球体からの出力の一部は前大脳の側角に投射する.
側角において湿,乾,冷糸球体からの投射を受ける領域は,小糸球体 (一般の匂い) や大糸球体 (性フェロモンの匂い) からの投射領域とは異なっているため,側角には触角で受容した種々の感覚情報を整然と並べて表現する「感覚地図」があると考えられる.
あるいはこれはむしろヒトの視床下部に見られる,引水行動,体温調節行動,摂食行動,性行動などの本能行動の中枢を整然と配置した「行動地図」のようなものなのかもしれない.
第7章 キノコ体は景色の記憶に関わる
キノコ体
デュジャルダンは昆虫のキノコ体を初めて顕微鏡で観察し,その複雑に折りたたまれた構造から哺乳類の大脳皮質を連想した.
キノコ体の機能は現在でも未知の部分が多いが,匂いなどの学習・記憶に関係していることが分かってきた.
ツチスガリの景色記憶
ティンバーゲンは実験により,ツチスガリ (狩りバチの1種) が巣の周りの景色の記憶を手掛かりに帰巣することを見出した.
ゴキブリの景色記憶とキノコ体
著者らは脳手術に強いワモンゴキブリを用いて,景色の記憶に関わる昆虫の脳領域を突き止めることを考えた.
そのためにはまず,ゴキブリに景色を記憶する能力があることを確かめる必要がある.
そこで著者らは床が高温 (約50℃) の円形の広場にゴキブリを投入し,ゴキブリが周りに配置された視覚的パターンを学習・記憶して,床のスポット状の熱くない部分 (ゴール) に早く到達できるようになるかを調べた.
その結果,ゴキブリにはゴールの周囲の景色を記憶する能力があることを確かめられた.
(実験はゴキブリが床の模様や床に残したフェロモンの匂い,壁の匂いを手掛かりにしている可能性を排除できるように工夫されている.)
次に著者らは脳を局所的に破壊したゴキブリを用いて同様の実験を行い,景色の記憶に障害が現れる脳領域を調べた.
実験では
- 学習行動の障害が視覚機能や運動機能の障害によるものではないことを保証するために,ゴールが見える対照実験を行った.
- ゴキブリの正常な探索行動を促すために,学習時に高温にさらされるゴキブリのストレスを減らす工夫を行った.
- 脳の破壊箇所を明確に特定できるように,脳の局所破壊はメスやハサミではなく,微小なアルミニウム薄片の埋め込みによって行った.
- キノコ体を切断したゴキブリの実験の対照実験として,頭部のクチクラ (外骨格) の一部を切り取って,そのまま元に戻した「無手術」のゴキブリと,キノコ体の側方にアルミ片を埋め込む「対照手術」を行ったゴキブリが用いられた.
実験の結果,キノコ体を切断したゴキブリに学習の障害が現れ,キノコ体が場所の記憶に関わることが示唆された.
またゴールが見える対照実験では,キノコ体を切除されたゴキブリもゴールにたどり着けたことから,場所の要素を含まない単純な視覚学習にはキノコ体は必要ないと考えられる.
行動中のキノコ体ニューロンの活動
著者はゴキブリの行動中にキノコ体のニューロンの活動を記録する技術を開発した.
そこでゴキブリの行動中のある神経活動を記録すると (記録はキノコ体の出力ニューロンからのものと推定された),ゴキブリが右にターンするときに,ターンの開始に0.5秒から2秒先行して活動電位の頻度が上昇することが分かった.
これは哺乳類の大脳皮質の運動前野・補足運動野における準備電位に類似しており,自発性運動の準備・企画などの高次の運動制御機能に関与するニューロンが昆虫のキノコ体に存在することを示唆している.
しかしキノコ体を破壊しても自発性運動は正常に見られることから,最終的に運動を決定・選択する脳領域は別にあると考えられる.
キノコ体出力ニューロンが投射する外側前大脳や内側前大脳が,そのような脳領域の有力な候補である.
キノコ体の内部構造
ゴキブリのキノコ体は,キノコの傘と柄に当たる部分と,柄から分岐したα葉とβ葉の4つの部分から成る (図10参照).
傘はキノコ体の入力部位であり,α葉とβ葉はキノコ体の出力部位である.
ケニオン細胞は傘からα葉とβ葉に信号を伝達する.
*
キノコ体はハチなどの社会性昆虫やゴキブリなどの群居性昆虫で発達しており,キノコ体を構成するケニオン細胞数は脳の全ニューロンの3~4割を占める.
*
キノコ体はスラブという明暗の薄層から構成され,暗スラブを構成するケニオン細胞は明スラブを構成するケニオン細胞よりも太い軸策を持つため,濃く見えることが分かった.
(これは年輪において細胞密度が大きい箇所が濃く見えるのと同じ事情である.)
またキノコ体にはあるスラブのみから信号を受け取る出力ニューロンがあることから,スラブは何らかの機能的なまとまりを反映した構造単位であると推測される.
このような構造単位は,哺乳類の大脳皮質にある機能コラムを想像させる.
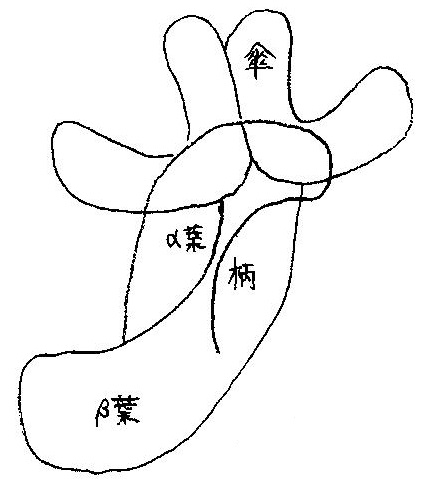
図10 ゴキブリのキノコ体
昆虫の脳の基本設計と記憶システム
キノコ体と他の脳領域との協調的な働きを考える上で,キノコ体を中心とする脳内の信号処理の流れを仮説的にまとめると,図11のように感覚中枢から胸部運動中枢に至る経路は,その長さに応じて4つの並列的な経路に分類できる.
第1の経路はゴキブリでは逃避行動の引き金を引くのに寄与し,第2の経路はハエでは飛翔制御に関係し,これらは素早い反射的な反応を担っていると言える.
第3の経路は配偶行動などの内的にプログラムされた行動に関与し,第4のキノコ体を経由する経路は匂いや場所を記憶し,経験に基づく行動を実現するのに必要とされると考えられる.
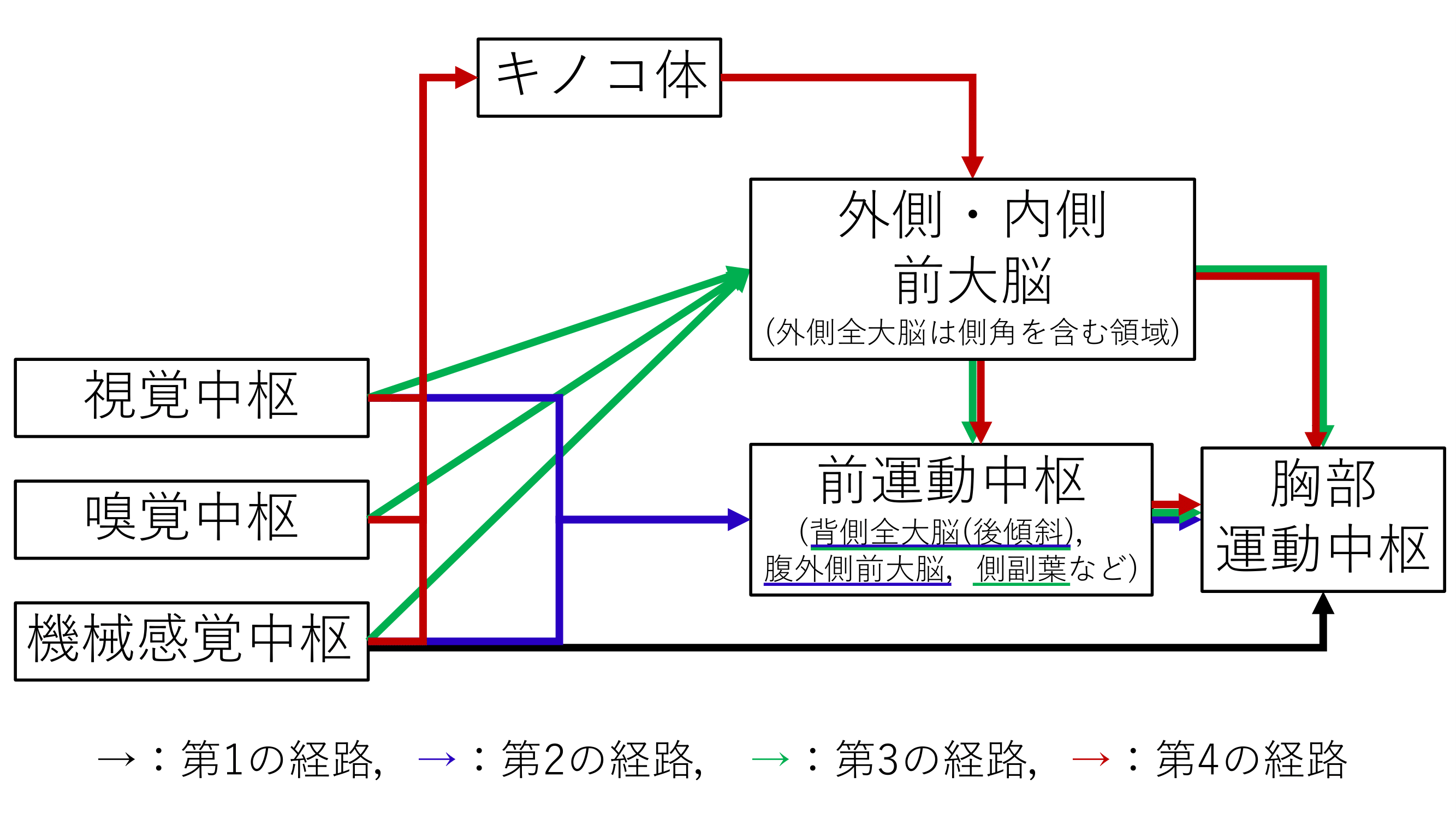
図11 キノコ体を中心とするゴキブリの脳における信号処理の仮説的な流れ
このような反射から高度な学習行動に至るまでの神経経路の重層的・並列的構造は,比較的単純な行動の神経経路の上に新しい経路が重層的に付け加わるような進化のプロセスを反映していると考えられ,哺乳類における
- は虫類脳 (反射脳)
- 哺乳類原脳[大脳辺縁系] (情動脳)
- 新哺乳類脳[大脳新皮質] (理性脳)
の三位一体構造 (図12参照) にも類似している.
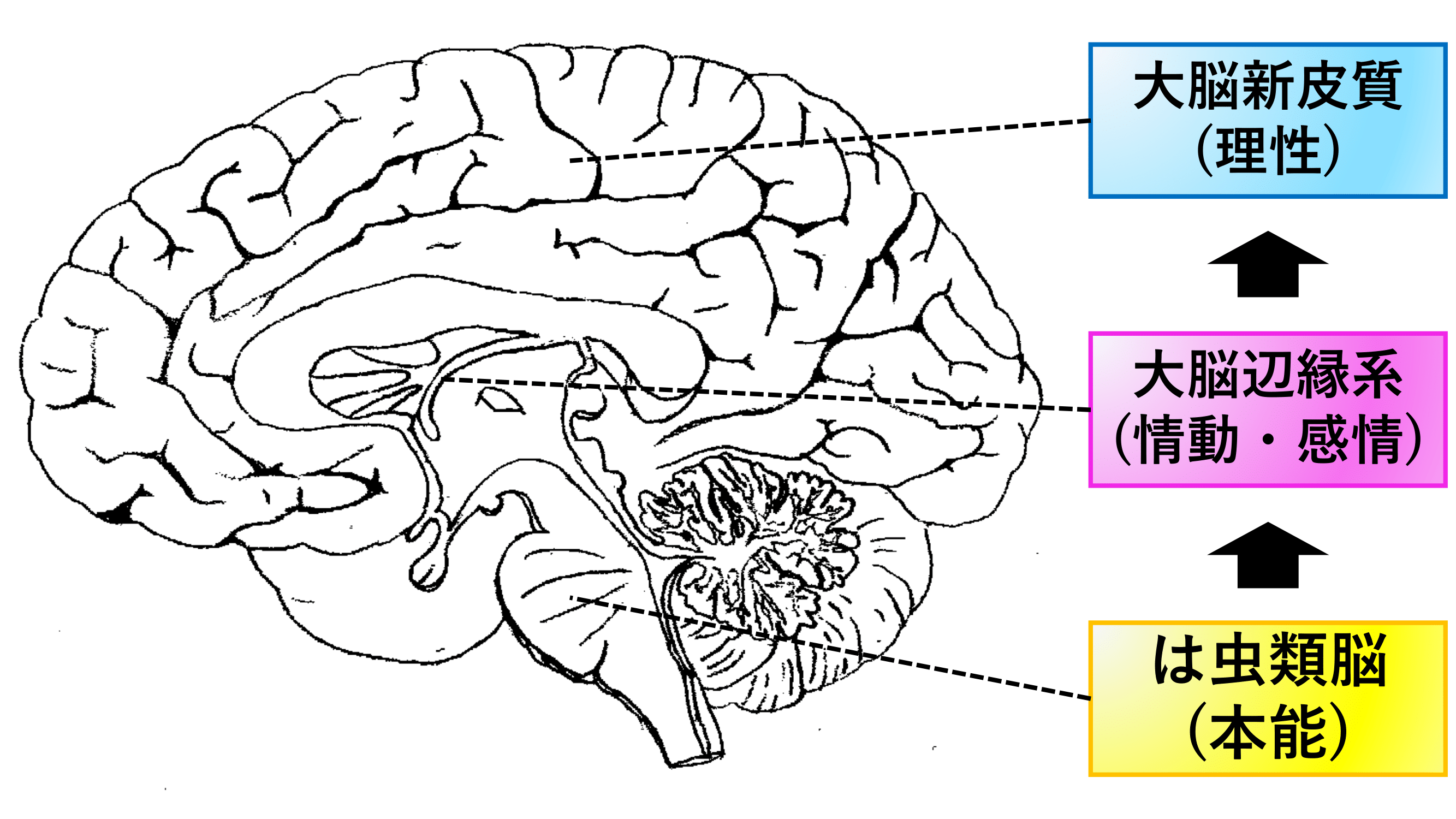
図12 三位一体脳
第8章 匂いの学習と記憶
匂いの学習と記憶
学習を「経験に基づく行動の変化」と定義し,学習された反応を保持する過程を「記憶」と呼ぶ.
従来の常識に反して,多くの昆虫が生存に密接に関わる局面において,優れた学習能力を示すことが分かってきた.
コオロギの学習
4日間絶水させたコオロギに匂いとともに水を与えたところ,水と連合させた匂いの嗜好性が上昇した.
このような連合学習は古典的条件付けと呼ばれ,今の場合,匂いを条件刺激,水を無条件刺激,匂い嗜好性の上昇を条件反応という.
- 1回の訓練 (水と匂いの対提示) でも学習は成立した.
- 十分な間隔を空けて訓練を繰り返すと,記憶が長く保持される傾向が見られた.(短期間の詰め込み学習では記憶が長持ちしないのは昆虫も同じである.)
- 学習のためには,条件刺激 (匂い) を無条件刺激 (水) に先行して与えることが必要であった.
記憶の三つの相について
学習訓練の20分程度後まで持続する記憶を短期記憶,1日後まで持続する記憶を長期記憶,短期記憶と長期記憶を結ぶ記憶を中期記憶と呼ぶ.
短期記憶なニューロンの持続的な電気的活動によって実現されるのに対し,長期記憶は生化学的な仕組みによるシナプス伝達効率の変化に加えて,タンパク質の合成による新たなシナプスの形成によるものと考えられる.
記憶の長期保持
コオロギの幼虫期にペパーミントの匂いを水 (または食塩水) と,バニラの匂いを食塩水 (または水) と連合学習させると,コオロギは水と連合した香りを嗜好するようになり,その記憶は成虫になってからほぼ生涯にわたって保持された.
(ここで水は報酬に,食塩水は罰に対応している.)
またコオロギが成虫になってから,水と食塩水と対提示する匂いを入れ替えた逆転訓練学習を行ったところ,コオロギは新たに水と連合学習させた匂いを嗜好するようになったことから,記憶は刷り込みと違って柔軟に書き換えられるものであることが分かる.
記憶容量
同様の実験により,コオロギは同時に7組の (14種類の) 匂いを記憶できることが確かめられた.
以上のコオロギをはじめとする昆虫の学習の速さ,記憶の長期にわたる保持,記憶の書き換えの柔軟性,記憶容量の大きさは,ラットやマウスなどの哺乳類の学習能力にも劣らない.
ミツバチの匂い学習に関わる脳の領域
匂いと砂糖水の条件付け (第6章) を行ったミツバチの脳を金属の針で局所的に冷却し,ニューロン活動を抑えたときに,条件付けの記憶に現れる影響を調べた.
すると触角葉は学習の約2分後に,キノコ体のα葉は学習の3~4分後に,キノコ体の傘は学習の約5分後に冷却すれば,記憶障害は現れなくなった.
このことからミツバチの匂いの記憶では,脳の複数の領域がそれぞれ異なる時間をかけて記憶を定着させていると考えられる.
ショウジョウバエの匂い学習の分子遺伝学
ショウジョウバエは遺伝学の実験材料に適している.
そこでまずショウジョウバエに匂いA (または匂いB) とともに電気ショックを与え,また電気ショックを与えない状態で匂いB (または匂いA) を流す学習訓練を行ったところ,通常のショウジョウバエは電気ショックと連合させた方の匂いを避けるようになった.
匂い学習ができない突然変異体
ところが匂い学習のできない突然変異体も見つかっている.
そのうち「ダンス」「ルタバガ」「DCO」と呼ばれる3種類の突然変異体は,サイクリックAMPを介したシグナル伝達系に関わる酵素タンパク質を作る遺伝子に異変があった (AMPはアデノシン-1-リン酸).
- ダンス
- 学習した記憶を数十分で消失(短期記憶の障害).サイクリックAMPの分解酵素に異常.
- これはサイクリックAMP濃度の慢性的な上昇をもたらし,サイクリックAMPシグナル伝達系の正常な働きを阻害.
- ルタバガ
- 短期記憶に障害.サイクリックAMPを合成するアデニル酸シクラーゼの1種に欠陥.
- アデニル酸シクラーゼは匂いと電気ショックの同時発生の検知に関係すると考えられているが,その詳しい仕組みは不明.
- DCO
- 中期記憶に障害.サイクリックAMP依存性タンパクキナーゼの触媒サブユニットの1つに欠陥.
- サイクリックAMP依存性タンパクキナーゼは,サイクリックAMPがあると活性化され,タンパク質をリン酸化する触媒として機能する.
- サイクリックAMP依存性タンパクキナーゼの活性化は,チャネルタンパク質のリン酸化による数時間程度のシナプス伝達効率の変化をもたらす.さらにクレブと呼ばれる分子を活性化させ,新たなシナプス形成に必要な遺伝子の転写を開始させ,長期記憶を促す.
以降,サイクリックAMP分解酵素,アデニル酸シクラーゼ,サイクリックAMP依存性タンパクキナーゼをコードする遺伝子をそれぞれダンス遺伝子,ルタバガ遺伝子,DCO遺伝子と呼ぼう.
ハエの匂い学習へのキノコ体の関与
特定の遺伝子が脳のどの組織で発現しているかをエンハンサートラップ法にて調べると,ダンス,ルタバガ,DCO遺伝子はキノコ体のケニオン細胞において高いレベルで発現していることが分かった.
またこれらの遺伝子がコードするタンパク質 (サイクリックAMP分解酵素,アデニル酸シクラーゼ,サイクリックAMP依存性タンパクキナーゼ) はいずれもキノコ体に高濃度で存在していることが分かった.
このことから条件付けをもたらす匂いと電気ショックとの連合過程はキノコ体で起こると考えられた.
*
GAL4エンハンサートラップ法にて,ルタバガ変異体のケニオン細胞においてのみルタバガ遺伝子を正常に発現させると,短期記憶は正常になった.
このことから短期記憶の形成にはケニオン細胞の正常な働きが不可欠であることが分かる.
*
Shibire (日本語の「しびれ」に由来) と呼ばれる突然変異体では,シナプス小胞の再取り込みに必要なタンパク質をコードしているダイナミン遺伝子に欠陥があり,この変異型遺伝子から作られるタンパク質は高温になると機能しなくなる.
この変異型遺伝子をケニオン細胞でのみ発現させ,高熱処理をしたときだけシナプス伝達が阻害されるような遺伝子組み換えハエを作成したところ,記憶の形成は阻害されず,記憶の読み出しに障害が現れた.
これは条件付けの記憶の形成が,ハエではケニオン細胞よりも上流で行われることを示している.
記憶の分子解剖──長期記憶における遺伝子転写調節タンパク質の関与
前述のクレブは遺伝子の転写を調節する働きを持つタンパク質 (転写調節因子) である.
- ハエのクレブを不活性化させると,間隔を空けた訓練の長期記憶が阻害された.
- ハエのクレブを常に活性化させると,1回の訓練で長期記憶が形成された.
記憶の形成を阻害する薬と促進する薬
昆虫の長期記憶形成の「分子スイッチ」として働くシグナル伝達経路は以下のように,前述のサイクリックAMPシグナル伝達系と,その上流のNO-サイクリックGMPシグナル伝達系を含むことが分かった.
- NO-サイクリックGMPシグナル伝達系
-
- NO合成酵素 (NOS) の働きにより,アルギニン (Arg) から一酸化窒素 (NO) が合成される.
- NOは細胞膜を気体として通過して周囲のニューロンに拡散し,水溶性グアニル酸シクラーゼ (sGC) という酵素を活性化させる.
- sGCがサイクリックGMP (cGMP) の合成を促進する.
- サイクリックAMPシグナル伝達系
-
- cGMPはCNGチャネルに結合してチャネルを開かせ,カルシウムイオン (
 ) を流入させる.
) を流入させる. - はカルモジュリンと呼ばれるタンパク質を活性化させる.
- 活性化したカルモジュリンはアデニル酸シクラーゼ (AC) を活性化させる.
- 活性化したACはATPからサイクリックAMP (cAMP) を合成する.
- cGMPはCNGチャネルに結合してチャネルを開かせ,カルシウムイオン (
↓
サイクリックAMP依存性タンパクキナーゼ (PKA) の活性化
↓
クレブの活性化
↓
長期記憶の形成
- サイクリックAMPシグナル伝達系とNO-サイクリックGMPシグナル伝達系のどちらが上流にあるかを調べるには,一方の阻害剤ともう一方の促進剤を投与する薬理学アプローチが有効である.
- カンデルはサイクリックAMPシグナル伝達系がマウス (哺乳類) とアメフラシ (軟体動物) に共通の長期記憶形成に関わるメカニズムであることを見出し,ノーベル賞を受賞した.
- 動物の記憶形成のメカニズムには,シグナル伝達系のこのような共通性だけでなく,多様性もある.
ミツバチの嗅覚学習に関わるニューロン
ミツバチの触角や吻に砂糖水をつけると,食道神経下節の中央に位置する腹側不対中央ニューロンが10秒以上にもわたって応答した.
また匂いを提示した後に,砂糖水の無条件刺激を与える代わりに不対中央ニューロンを活性化させても,匂いを吻伸展反応に条件付けることができた.
不対中央ニューロンを活性化させただけでは吻伸展反応は起こらなかったことから,不対中央ニューロンは無条件刺激の報酬にのみ関わっていると考えられる.
不対中央ニューロンの神経伝達物質はオクトパミンであると考えられている.
そこで不対中央ニューロンを活性化させる代わりに,それが投射する触角葉とキノコ体の傘に個々にオクトパミンを注射すると匂いと吻伸展反応の条件付けが起こったことから,不対中央ニューロンの神経伝達物質であるオクトパミンがミツバチの脳において,報酬による条件付けを誘導することが示唆された.
昆虫の学習における罰系と報酬系
さらなる研究により,学習の種類 (嗅覚学習か視覚学習か) や報酬の種類 (砂糖水か水か) や罰の種類 (電気ショックか食塩水か),種の違い (ミツバチかハエかコオロギか) によらず,一般に昆虫の学習においてオクトパミン作動性ニューロンが報酬情報を,ドーパミン作動性ニューロンが罰情報を伝達していることが示唆された.
これは脊椎動物において,ドーパミン作動性ニューロンが報酬情報を伝えているのとは対照的である.
さらに高次の匂い学習
昼間は匂いA (またはB) と水,匂いB (またはA) と食塩水を連合させ,夜間は逆に匂いB (またはA) と水,匂いA (またはB) と食塩水を連合させる訓練を行ったコオロギは,昼か夜かというコンテキストに応じて水と連合させた匂いを選ぶことができた.
(匂いA,Bとしてはペパーミントの香りとバニラの香りが用いられている.)
ワモンゴキブリにも同様の能力がある.
このような高次の匂い学習能力に関わる昆虫の脳の仕組みの解明は今後の課題である.
第9章 ミツバチのダンス
本章ではミツバチの8の字ダンス (尻振りダンス) を取り上げる.
8の字ダンスを実現する脳の仕組みはまだ分かっていないが,解明に向けた準備が整いつつある.
ミツバチの餌探し飛行
ミツバチの効率の良い採餌には,よい蜜源を見つけた働きバチが仲間のハチに,ダンスによって蜜源のありかを伝えるコミュニケーション能力が大きく貢献している.
ハチの社会と超個体
省略する.
8の字ダンスの信号
- 円形ダンス
- 餌が巣の近くにあることのみを伝える.
- 8の字ダンス (尻振りダンス)
- 遠くにある餌場を伝える.
- 8の字の中央における直進方向が餌場の方向に関係し,直進方向の鉛直上向きとの成す角が,巣から見た餌場の方向の太陽方向との成す角に等しい (図13参照).すなわち鉛直上向きを太陽の方向に見立てて,餌場の方向を表している.
- 直進部で翅を震わせて出す音の持続時間が餌場までの距離を表す (1秒が約200mに対応).
暗い巣箱の中では,ミツバチは仲間のダンスを視覚的にではなく,触角を用いて音として読み取る.
ダンスは蜜源だけでなく,水や花粉のありか,新しい巣の候補地の伝達にも転用される.

図13 8の字ダンスでの方向指示
ミツバチ・ロボットによる検証
当初は餌場から戻ったミツバチはダンスによってではなく,身に付けてきた花の香りや吐き出した蜜によって餌場の情報を伝えているのではないかという反論があった (ダンスをするミツバチはしばしば蜜を吐き戻して周囲のハチに伝える).
そこでダンスを踊るロボットバチを用いた検証実験を行ったところ,ロボットがダンスによって示した方向の餌場に多くの働きバチが向かった.
これによりミツバチのダンスによる情報伝達が証明された.
またロボットを用いた検証実験により,ダンスが機能するには翅を震わせる音が必要であること,蜜を吐き戻して周囲のハチにサンプルとして与える必要があることが確かめられた.
方向と距離をどのようにして測るのか
太陽の見えない閉鎖的な場所では,ミツバチは太陽の方向を鉛直上向きに読み替えてダンスを踊るが,空が見えるようにした巣では直接餌場に向けてダンスを踊る.
*
従来,ミツバチは巣に帰る際,消費したエネルギー量 (蜜の量) で巣までの移動距離を見積もっていると考えられていた.
ところが餌場までの通路の壁に,通路と (したがってミツバチの飛行方向と) 平行または垂直な縞模様を描いておくと,ミツバチは垂直な縞模様の場合には通路上のほぼ正しい餌場の位置に戻ってくることができた.
このことからミツバチは移動時のオプティカルフロー (景色の流れ) を基に移動距離を見積もっていると考えられる.
また通路に順風・逆風を流した場合にもミツバチは正しい位置に戻ってきたため,エネルギー消費量は距離の計測に関係していないと考えられる.
さらに本来は円形ダンスをするような巣の近くに,オプティカルフローをもたらす縞模様を施した通路の餌場を置くと,ミツバチに餌は遠くにあると誤認させ,8の字ダンスを躍らせることができた.
ダンスの進化
リンダウアーは4種類のミツバチの進化した順を,ダンスの形態から次のように推測した.
- ヒメミツバチ (野外の水平面でのダンス,直接餌場の方向に向けてダンス)
- オオミツバチ (野外の垂直面でのダンス,太陽方向を鉛直上向きに変換)
- トウヨウミツバチ,セイヨウミツバチ (閉鎖空間の暗闇での垂直面でのダンス,太陽方向を鉛直上向きに変換)
【コメント】
もっともらしいが,2と3は逆でも良いように思われる.
閉鎖空間の暗闇で太陽方向を鉛直上向きに変換するダンスを獲得した種 (3) が,再び巣版を野外に持つ種に進化したが,ダンスの形態には暗闇でのダンスの名残がある (2) という可能性も考えられる.
進化の順序を推測するには,ダンスの形態だけでなく遺伝子の相違などを総合的に考える必要がある.
ミツバチは陳述記憶をもつ
ヒトの記憶は陳述記憶と手続き記憶に分類される.
- 陳述記憶:意図的に想起でき,記憶の内容を言語で表現できる.海馬などが関係.
- 手続き記憶:「体が覚えた」記憶で,意図的に想起できず,動作の実行中も想起していることが意識されない.小脳などが関係.
餌のありかについての働きバチの記憶はダンスによって「陳述」されるため,陳述記憶と見なせる点でユニークである.
【コメント】
ただしここでの「陳述」は擬人的な表現であり,実際の昆虫の行動はむしろ機械的・無意識的・自動的なものである可能性がある.
(これまで繰り返し用いられてきた,昆虫が匂いを「識別する」というような表現も同様である.)
これは昆虫に限らず,ヒトが記憶を想起する場合にもある程度当てはまるかもしれない.
第10章 ハチやアリの偏光コンパス
太陽コンパスと偏光コンパス
景色を記憶することが困難な長距離にわたる移動の際には,昆虫は太陽の位置をコンパスとして利用する.
そのためには昆虫は1日における時刻を分かっており,かつその時刻における太陽の位置を知ってる必要があるが,実際,昆虫にはそのような能力がある.
さらにアリやハチは空の偏光パターンをコンパスに用いることができ,直接太陽が見えなくても青空の一部が見えさえすれば巣に帰ることができる.
サバクアリの偏光コンパス
サバクアリはフェロモンの道しるべを残さないにも関わらず,周りの景色に目印となるものがない状況下でも真っ直ぐに巣に帰ることができる.
これはアリが,常に自分の位置の巣からの変位の情報を保持しているためであると考えられる.
[巣からの変位ベクトルは「理論的には,巣から歩いてきたすべての距離と,曲がってきたすべての角度を積算することで計算できる」ことについて,変位は  である.]
である.]
実際巣Nから餌場Fにやってきたアリを,周りの景色が見えない状態で他の位置に移動させると,巣に戻ろうとしてその点から変位の逆ベクトル  だけ直進した.
だけ直進した.
*
太陽コンパスと偏光コンパスの情報が矛盾する場合,アリは偏光コンパスを優先的に用いた.
太陽コンパスはアリにとって,偏光コンパスを使えないときのバックアップシステムである.
空の偏光パターン
大気に散乱された光の偏光面は,散乱点と太陽と観測者を含む面に垂直であり (レイリー散乱の法則),このため天球上の散乱点に対して偏光方向は,太陽を中心として同心円状に分布する (図14参照).
偏光の強度は太陽から90度離れた大円上で最大となる.
よってどの時刻でも偏光パターンは(反)太陽子午線に関して対称であり,太陽はこの対称線上で長波長の光に富む側にある.
こうしてアリやハチは太陽の方向を把握する.

図14 空の偏光パターンと太陽の位置
内的鋳型と走査モデル
ハチやアリは実際の空の偏光パターンの近似的な内部鋳型を持ち,これを実際の空の偏光パターンと比較することで帰巣の方向を定めている.
偏光情報の受容と処理の神経機構
- ハチやアリは複眼の背縁部にある紫外線受容細胞で空の偏光を検知している.
- 偏光受容領域における紫外線受容細胞の感桿の微絨毛は,互いに直交する2方向に配列されており,それぞれ90度異なる方向の偏光の強さを検出する.
- 偏光受容領域における紫外線受容細胞では微絨毛に沿って棒状の光受容分子が配列されており,微絨毛と平行な方向の偏光に感度良く反応する.
*
偏光受容領域において空の各方向に向けられた紫外線受容細胞の微絨毛の配列分布は,昆虫の偏光パターンの内部鋳型とおおよそ一致しており,ウェーナーはこの微絨毛の配列パターンと空の偏光パターンが最も良く一致する方向に体を向けることで,昆虫は(反)太陽子午線の方向を検出しているという「走査モデル」を唱えた.
*
コオロギの偏光受容領域のメダラにある介在ニューロンは,ある方向の偏光に対して興奮性の反応を示し,それと90度異なる方向で抑制性の反応を示した.
これはある方向の微絨毛の紫外線受容細胞による興奮性の反応と,微絨毛の方向が90度異なる紫外線受容細胞による抑制性の反応が拮抗していることから理解できる.
[ある方向の偏光に対しては興奮性の反応が勝ち,それと90度異なる偏光に対しては抑制性の反応が勝ち,その中間の方向の偏光に対しては興奮と抑制が相殺される傾向にある,というように.]
走査モデルの検証
ミツバチに偏光板を通した紫外線を当てると,光強度が極大となる方向を(反)太陽子午線の方向と勘違いして8の字ダンスを踊ったことから,走査モデルが検証された.
中心複合体は偏光コンパスの座か
偏光情報は
メダラ → 前大脳の前視結節 → 側副葉 → 中心複合体
と伝えられる.
中心複合体と側副葉を偏光コンパスに基づく定位行動の中枢とする仮説があり,中心複合体にあるコラム構造が昆虫の定位行動とどのように関わっているかに興味が持たれる.
ハチの空間地図
巣箱と餌場を何度か往復させたミツバチを捕獲し,周りが見えない状態で巣の周囲の任意の位置まで持ち運び放すと,一部のミツバチは巣が見つからないため,一度餌場に戻ってから帰巣した.
このように巣箱と餌場の間のルート周辺の任意の点から餌場に戻れることは,ミツバチがヒトと同様に2次元的な空間地図を有していることを示唆している.
第11章 微小脳と巨大脳
進化の2つの頂点
ヒトと昆虫はそれぞれ新口動物と旧口動物の頂点に君臨する (図15参照).
(エボ・デボ生物学により,例えばボディ・プランを決める遺伝子は両者に共通していることが分かった.)
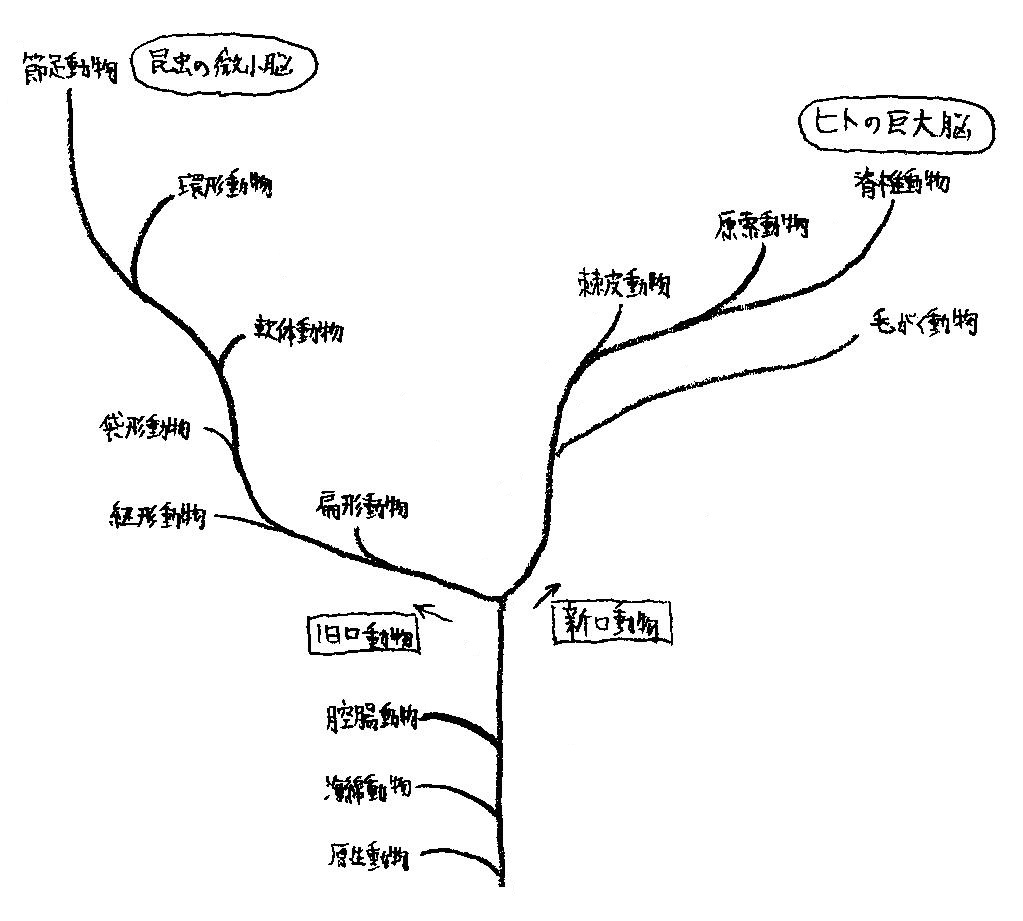
図15 動物の系統と脳の進化
ピアンカの理論
ピアンカは生物を繁殖戦略に応じてr戦略者とK戦略者に分類できるとし,昆虫をはじめとする短命で小型な陸上無脊椎動物はr戦略者,ヒトをはじめとする長命で大型な陸上脊椎動物はK戦略者と見なせるとした (rやKは成長曲線のパラメーターであり,ここではその意味は重要でない).
- r戦略者
-
- 不安定な環境に生息,体が小さい,世代時間が短い,個体数が多い,多産
- 短期間に多くの変異体を生み出すことで環境の変化に対応
↔ 微小脳による鋭い感覚能力,素早い運動能力
- K戦略者
-
- 安定な環境に生息,体が大きい,世代時間が長い,個体数が少ない,少産
- 長い生涯にわたり学習によって行動を変化させて環境の変化に対応
↔ 巨大脳による高度な学習能力
ピアンカの理論は昆虫と哺乳類の違いを理解するのに役立つけれど,短期間に多くの変異体を生み出すことで環境の変化に対応する昆虫にも高度な学習能力が備わっていることを説明できない.
そこでこれを補う考え方として,次にマイヤの考察を紹介する.
マイヤの考察
「食われない」ための行動 (外的からの回避行動) や「子孫を残す」行動は,素早さまたは確実性が要求されるため,基本的には遺伝的にプログラムされた生得的な定型行動が適している.
ところが「食っていく」ための行動に関しては,適切な行動を遺伝的にプログラムするには限界があり,哺乳類だけでなく昆虫においても学習能力が重要となる.
このようにマイヤの考察は,昆虫と哺乳類の共通性を理解するのに役立つ.
【コメント】
分からなくもないが,これはやや詭弁であるようにも思われる.
「食う」行動においても,目の前の動くものを餌と判断して食らいつくような生得的な行動がまず重要であると考えられなくもない.
微小脳という視点から
著者は微小脳を「外骨格を持つ動物の脳」と定義した.
昆虫のように外骨格を持つ動物は,力学的に大きくなれないという制約がある (大きくなると,体を支えるのに必要な分厚い外骨格の重みによって身動きが取れなくなる).
また昆虫の気管呼吸方式は体が大きくなると効率が悪くなる (酸素濃度の高かった古生代の,80cm程度の巨大トンボあたりが限界).
昆虫は小さいがゆえに,小さくなると体温を維持するためのエネルギー効率が悪くなり,小さくなれない恒温動物との直接的な競争を避けられた.
[体が小さくなると体積当たりの表面積が小さくなり,体温の維持が困難になること自体は,変温動物の昆虫も同じである (p.149).]
微小脳と巨大脳の比較
- ヒトの巨大脳は膨大な情報の精密な処理に適しているのに対し,昆虫の微小脳は情報を取捨選択する「粗い情報処理」による「速い情報処理」に適している.
- ヒトの巨大脳では大脳皮質のコラム構造のように多数のニューロンの集団が機能単位となっているのに対し,昆虫の微小脳ではキノコ体などでの多数のニューロン集団による情報処理と,昆虫に典型的な少数のニューロンによる制御が巧みに連携をとっていると考えられる.
- 昆虫の脳の機能を少数のニューロンの働きに還元して理解できる例:飛翔系の「偏差検知ニューロン」,匂い学習に関わる「不対中央ニューロン」
- ヒトの巨大脳に比べて昆虫の微小脳では,階層 (直列処理の連なり) が非常に浅い.
- ヒトの巨大脳では感覚情報はあまり加工を受けずに感覚野に投射され,視覚系では顔の表情のような像の細かい特徴まで識別できるのに対し,昆虫の微小脳では同様の視野再現構造がロビュラプレートに見られるものの,情報の再現は粗く,また像の形態よりも運動の情報が重視・抽出される.
- ヒトの巨大脳では側頭葉や小脳などに学習行動の膨大な記憶を貯蔵できるのに対し,昆虫では行動の大部分が遺伝的に決められている.
- ヒトの巨大脳は複数のニューロンにより複雑で精密な運動を実現するのに対し,昆虫の微小脳では少数のニューロンによる素早い制御を実現する.
ヒトと昆虫
省略する.